Life History & Behaviour
Feeding
As a result of being benthic dwellers that feed on algae and seaweed attached to the sea floor (Miskelly 1968), the sea urchin mouth is positioned orally towards the substratum (Fenner 1973; Ruppert et al. 2004). They feed by scraping matter off the surface of substratum with their five strong teeth (Nichols 1967) that are a part of their highly developed protrusible jaw apparatus, the Aristotle’s lantern (Ruppert et al. 2004). These teeth are projected off five large calcareous plates (Ruppert et al. 2004). The Aristotle’s lantern consists of 40 skeletal pieces, bound with muscles and tendons, which control the teeth (Nichols 1967). There are three sets of muscles involved in controlling the teeth; these are the interalveolars, adductors and abductors (Nichols 1967). There are also specialised muscles for controlling the lantern; these are involved in protruding or retracting it or moving it from side to side (Ruppert et al. 2004). There is large variability seen in the diet of echinoids, while their major food source is algae, they also feed on a variety of plant and animal materials (Ruppert et al. 2004). Their versatile diet is due, in part, to the advanced grazing ability provided by the Aristotle’s lantern.
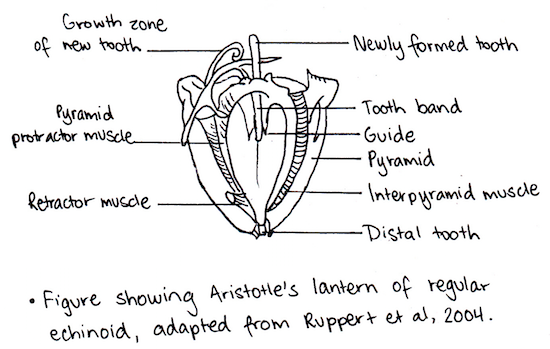
Locomotion (Information summarised from Ruppert et al. 2004)
Sea urchin locomotion is mainly controlled by tube feet and spines on the oral surface of the test. This has allowed them to succeed in both hard and soft substrates. By the combined use of their tube feet and spines, sea urchins can flip themselves back onto their oral surface if overturned. Their structures allow them to move in any direction without turning, as any ambulacral area can lead inlocomotion. Movement of the tube feet is achieved by the pumping of water into and out of the tube feet. A more detailed description of locomotion using the tube feet is presented under Anatomy & Physiology.
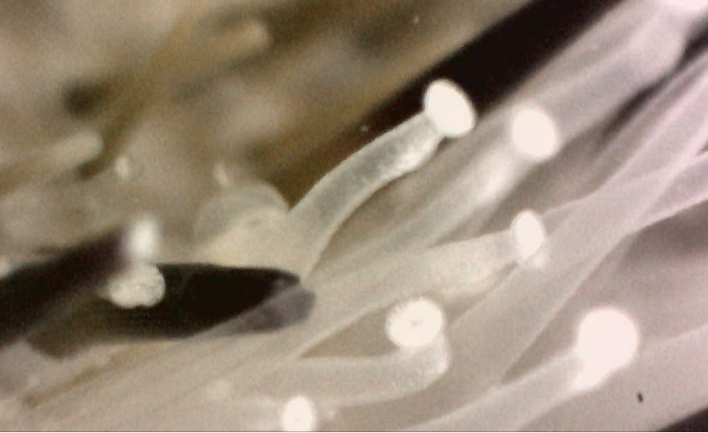 |
Temnopleurus alexandri tube feet extended
Photo: Monique Parisi |
Respiration
Echinoids have specialized structures for respiration; these are peristomial gills and tube feet (Ruppert et al. 2004). Generally, non-locomotor podia on the aboral surface are involved in respiration (Fenner 1973; Leddy & Johnson 2000). Echinoids that burrow in relatively anaerobic substrate have the most highly adapted respiratory podia (Fenner1973), as T. alexandri don’t do this, it can be assumed that they wouldn’t have these highly adapted respiratory podia.
Reproduction
T. alexandri are described as gonochoric, which means they have separate sexes (male and female individuals) (Ruppert et al. 2004). As a ‘regular’ urchin, they have five gonads, which are arranged internally along interambulacral plates (Ruppert et al. 2004). T. alexandri are broadcast spawners, releasing their eggs and sperm into the water column, where fertilisation occurs. The release of their gametes is controlled by a number of cues (Williamson & Steinberg 2002). Timing of gamete release is important because sperm only survives in the water column fora certain amount of time. Echinoids are reproductively successful because they release large amounts of gametes into the water column at one time, which can result in high-density populations (Williamson & Steinberg 2002).
Development
As deuterostomes, echinoderms display radial cleavage during development (Ruppert et al. 2004). Their coelomic cavities have entercoelous origin, which refers to the process of coelom and mesoderm formation from outpouching of the endoderm (Ruppert et al. 2004). The mouth forms from secondary invagination of the ectoderm and the anus from the blastopore. Metamorphosis differs between classes within the phylum; however, the anus in echinoids opens from the first plate to form in metamorphosis (Nichols 1967). Five plates surround the anal plate, which allows the formation of a strong test (Nichols 1967). During development, each skeletal element of a sea urchin is composed of a single calcite crystal (Nichols 1967). This is a unique characteristic of individuals within the phylum Echinodermata.
|